Fundamentals of Sound |
|
Fundamentals of Sound |
|
|
THE EAR AS A TRANSDUCER - Introduction
/ Outer & Middle Ear
|
|
|
(source) |
Transducers: Devices that convert
|
|
|
|
TRANSDUCTION PROCESS IN THE OUTER AND MIDDLE EAR |
|
|
|
|
|

Simplified graph of the ear: in anatomical context (left) and magnified & sectioned (right) |
 |
|
Schematic:
Longitudinal waves reaching the eardrum
(enlarge - animation) |
|
|
|
|
|
|
(a) (b) (c) (d) (a)
Eardrum motion
& (b)
Ossicle motion animations
|
|
|
TRANSDUCTION PROCESS IN THE INNER EAR
|
|
|
|
|
|
|
←---------------|
Top & Middle: two simplified
schematics of a
stretched-out cochlea; |
|
Above Left: Microphotographs of two intact cochleae
(top and middle)
and of a dissected one (bottom). |
|
|
Schematic cross-sections of the cochleae cochlea Illustrations of the three key compartments or scalas (scala: ladder) and the main transduction element: the Organ of Corti. |
Schematic close-up of the Organ of Corti
The Organ of Corti (OofC) is bounded |
|
|
Inner-Ear Innervation | |

Healthy (top) & damaged (below) hair cells
|
|
|
Videos outlining the auditory transduction process |
|||
|
|
|
|
| Additional Videos @ https://www.interactive-biology.com/physiologyvideos/ (videos 036-040) |
|||
|
|||
THE BASILAR MEMBRANE
|
|||
Basilar membrane (BM): A collection of interconnected, weakly-coupled, flexible fibers located at the basis of the Organ of Corti, in the inner ear (cochlea). |
250Hz:
1kHz:
4kHz: |

|
|
|
|
|
||
|
|
||
|
||
|
Conversely, and as already stated, critical bandwidth can be defined as the minimum frequency separation necessary for two ~equally strong, simultaneous sine waves to sound clearly apart, free from beating and/or roughness. Both, the beating and roughness sensations are perceptual attributes of amplitude fluctuation resulting from sound wave interference (discussed previously). Psycho-physiologically, the beating and roughness sensations are linked to:
[We will return to beating and roughness, when discussing musical timbre, consonance, and dissonance.] |
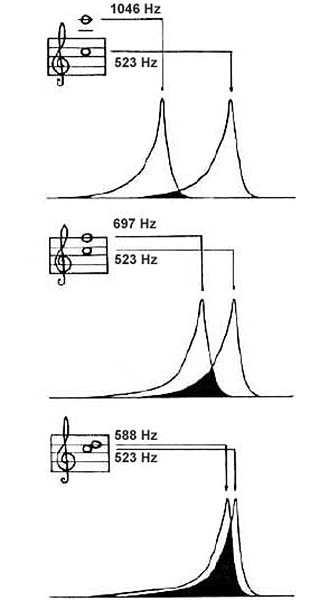 As the interval between two tones
of comparable levels decreases, their respective
disturbances on the basilar membrane (critical bands) increasingly
overlap, resulting in the sensations of roughness and beating |

The first 12 components of C3, shown as black circles
on a stretched music-notation 'stave'. |
| ||
|
|
|
Simultaneous Masking Term describing the ability of one tone or band of noise (masker) to cover or raise the audibility threshold of a second tone (signal). When two tones (or a band of noise and a tone) close in frequency are
presented simultaneously, one (masker) may mask or �cover� the other
(signal) depending on their level difference: the more intense tone may
mask the less intense tone. The response of the BM around the characteristic (resonant) frequency
is asymmetrical: it is larger above than below characteristic frequency.
Low frequency
tones are more likely to mask (i.e. are more efficient maskers than) high frequency tones. Simultaneous masking may be due to
|
 
As the level of a signal increases, the BM response
increases in magnitude and width (inverted v-shaped lines,
above). This means that stronger signals are able to cover
increasingly stronger simultaneous signals and increasingly
further removed in frequency. |
|
Temporal Masking During Forward Masking, a signal masks a tone that comes 0-200ms after it. Forward masking does not produce the broad masking effects of simultaneous masking.
During Backward Masking, a signal masks a tone that came
0-50ms before it.
|
|
|
|
|
Distortion
Suppression |
|
|
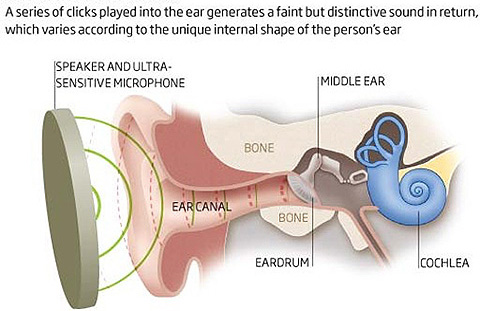
Otoacoustic emissions The ear acts not only as a microphone, receiving sound, but also as a speaker, emitting a series of tones referred to as otoacoustic emissions (OAEs). OAEs are classified depending on the context of emission (i.e. spontaneous vs. evoked and, if evoked, by what).
Watch this video describing an application of OAEs to hearing assessment and headset tuning. Here is another relevant product. |
 |
|
What Types of Hair Cells and Nerve Fibers Populate the Inner Ear? |
|
Video overview of the hearing transduction path and process (Dr. G. Bhanu Prakash - Animated Medical Videos) | |
|
| |



 (a) (b) (c) (d) (a)
Basilar membrane vibration animations and (b)
in-vivo (alive) vs. in-vitro (dead) vibration
measurements
|
|
|
(a) (b)
Outer hair-cell
(a)
motility,
and (b)
explanation,
of inner and outer hair-cell function | |
|
Temporal Coding | |
 |
Phase Locking and Rectification Neural response (neural firing) follows (or appears to be locked to) the positive peaks in the stimulus, firing only when the stereocilia are sheared in one direction.
This results in the neural signals of sinusoidal inputs
|
|
The process of phase locking is closely related to hearing's "temporal coding theory" of encoding frequency.
| |
|
Place Coding | |
|
| |
|
The above figure illustrates an alternative to the "temporal coding theory" of encoding frequency, referred to as "place coding theory."
As we've discussed, the basilar membrane (BM) responds
at different places for different frequencies, due to
its mass-stiffness gradient. | |
 |
The image to the left is a schematic diagram illustrating the signals sent to the brain when the basilar membrane is vibrating in response to a complex wave with many sine components. Each low-frequency component
sends individual signals since, as indicated above, the
frequency separation between the low-frequency components is larger
than the critical bandwidth. |
|
Overview Resources on Ear Anatomy and Function |
|
Dangerous Decibels from the National Institute of Health website. |
 |
Well-designed and concise overview of the anatomy and function of the ear, Part of NeurOreille's Journey Into the World of Hearing. |
 |
The Auditory System: Structure and Function, part of the Neuroscience e-Textbook site at the Department of Neurobiology and Anatomy of the McGovern Medical School, University of Texas. |
 |
|
|
|
|
|
|
|
|
Loyola Marymount University - School of Film & Television




















 ;
; 






